| Главная | Генетические алгоритмы | Нейросети | AI | Библиотека | Форум | Проекты | Поиск |

![]() Биологический нейрон
Биологический нейрон
Моделирование биологических нейронных сетей обоснованно и перспективно. Но для исследования НС необходимо иметь математическую модель биологического нейрона и биологической нейронной сети.
Центральная нервная система имеет клеточное строение. Единица – нервна клетка, нейрон. Нейрон имеет следующие основные свойства:
- Участвует в обмене веществ и рассеивает энергию. Меняет внутреннее состояние с течением времени, реагирует на входные сигналы и формирует выходные воздействия и поэтому является активной динамической системой.
- Имеет множество синапсов – контактов для передачи информации.
- Нейрон взаимодействует путем обмена электрохимическими сигналами двух видов: электротоническими (с затуханием) и нервными импульсами (спайками), распространяющимися без затухания.
Существуют два подхода к созданию искусственных нейронных сетей. Информационный подход: безразлично, какие механизмы лежат в основе работы искусственных нейронных сетей, важно лишь, чтобы при решении задач информационные процессы в НС были подобны биологическим. Биологический: при моделировании важно полное биоподобие, и необходимо детально изучать работу биологического нейрона.
Крупные работы в исследованиях биологических нейронных сетей принадлежат Эндрю Хаксли, Алану Ходжкину, Бернарду Катцу, Джону Экклзу, Стивену Куффлеру.
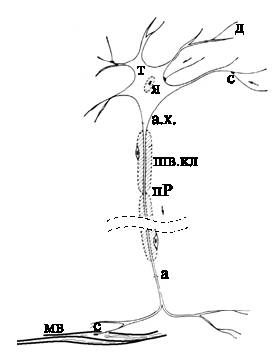 |
| (Рис.1) Биологический нейрон. |
Биологический нейрон содержит следующие структурные единицы:
- Тело клетки(т) — сома: содержит ядро (я), митохондрии (обеспечивают клетку энергией), другие органеллы, поддерживающие жизнедеятельность клетки.
- Дендриты (д) – входные волокна, собирают информацию от других нейронов. Активность в дендритах меняется плавно. Длина их обычно не больше 1 мм.
- Мембрана – поддерживает постоянный состав цитоплазмы внутри клетки, обеспечивает проведение нервных импульсов.
- Цитоплазма — внутренняя среда клетки. Отличается концентрацией ионов K+, Na+, Ca++ и других веществ по сравнению с внеклеточной средой.
- Аксон (а), один или ни одного у каждой клетки, – длинное, иногда больше метра, выходное нервное волокно клетки. Импульс генерируется в аксонном холмике (а.х.). Аксон обеспечивает проведение импульса и передачу воздействия на другие нейроны или мышечные волокна (мв). Ближе к концу аксон часто ветвится.
- Синапс (с) – место контакта нервных волокон — передает возбуждение от клетки к клетке. Передача через синапс почти всегда однонаправленная. Различают пресинаптические и постсинаптические клетки — по направлению передачи импульса.
- Шванновские клетки (шв.кл). Специфические клетки, почти целиком состоящие из миелина, органического изолирующего вещества. Плотно "обматывают" нервное волокно 250 слоями миелина. Неизолированные места нервного волокна между шванновскими клетками называются перехватами Ранвье (пР). За счет миелиновой изоляции скорость распространения нервных импульсов возрастает в 5-10 раз и уменьшаются затраты энергии на проведение импульсов. Миелинизированные волокна встречаются только у высших животных.
В центральной нервной системе человека насчитывается от 100 до 1000 типов нервных клеток, в зависимости выбранной степени детализации. Они отличаются картиной дендритов, наличием и длиной аксона и распределением синапсов около клетки.
Клетки сильно связаны между собой. У нейрона может быть больше 1000 синапсов. Близкие по функциям клетки образуют скопления, шаровидные или параллельные слоистые. В мозгу выделены сотни скоплений. Кора головного мозга – тоже скопление. Толщина коры — 2 мм, площадь — около квадратного фута.
![]() Нервный импульс
Нервный импульс
Нервный импульс (спайк) – процесс распространения возбуждения по аксону от тела клетки (аксонного холмика) до окончания аксона. Это основная единица информации, передаваемая по волокну, поэтому модель генерации и распространения нервных импульсов (НИ) — одна из важнейших в теории НС.
Импульсы по волокну передаются в виде скачков потенциала внутриклеточной среды по отношению к внешней среде, окружающей клетку. Скорость передачи – от 1 до 100 м/с. Для миелинизированных волокон скорость передачи примерно в 5 – 10 раз выше, чем для немиелинизированных.
При распространении форма спайка не меняется. Импульсы не затухают. Форма спайка фиксирована, определяется свойствами волокна и не зависит от того, каким способом создан импульс.
На рис.2 показаны нервные импульсы, возникающие в зрительном нервном волокне при воздействии вспышки света постоянной длительности для различных интенсивностей вспышки. Свет воздействовал на фоторецепторы и вызывал импульсацию в соответствующем зрительном волокне.
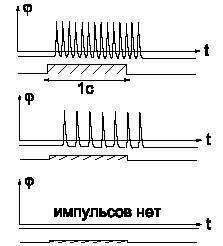 |
| (Рис.2) Потенциал цитоплазмы относительно межклеточной среды в зрительном волокне при воздействии световой вспышки различной интенсивности. |
Видно, что от интенсивности света зависит не амплитуда импульсов и их форма, а плотность и общее количество.
Для возбуждения и регистрации НИ можно использовать такую схему:
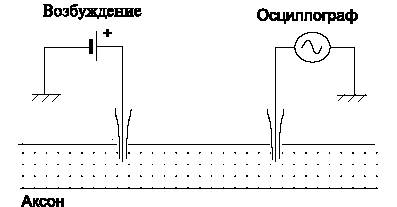 |
| (Рис.3) Возбуждение и регистрация нервных импульсов. |
Подавая на возбуждающий электрод электрические импульсы различной длительности и амплитуды, можно фиксировать возникающие при этом импульсы.
Оказалось, что зависимость минимального
возбуждающего тока, при котором возникают нервные импульсы, от времени
возбуждения имеет вид гиперболы: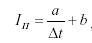
где
Iп - минимальный ток, при котором возникает импульсация
при данной длительности delete t возбуждения. Данная
формула - эмпирическа.
Параметры a и
b имеют следующий смысл. b называется
реобазой и задает минимальной ток возбуждения, при котором вообще
может возникнуть импульсация. Если возбуждающий ток I меньше
b, то импульсов не возникнет при любом delta t.
Количество электричества, необходимое дл возбуждения импульсов, при малых
delta t примерно постоянно: 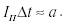
Эксперементально было открыто свойство рефрактерности: после того, как по волокну прошел нервный импульс, в течении несколько миллисекунд новый импульс не возбуждаются при любом Iп. По этому между нервным ипульсам всегда есть минимальный интервал времени - период реврактерности.
![]() Мембрана. Мембранный потенциал.
Мембрана. Мембранный потенциал.
Клеточная мембрана создает и поддерживает постоянную концентрацию веществ внутри клетки, механическую прочность клетки и транспортировку молекул и ионов в обоих направлениях. Для нервных волокон она обеспечивает проведение нервных импульсов по волокну.
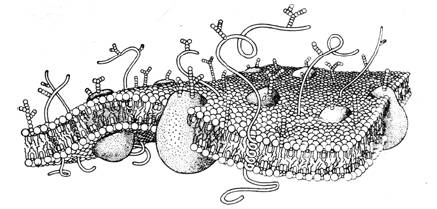 |
| (Рис.4) Клеточная мембрана. |
Мембрана состоит из двух слоев молекул липидов (жиров). Молекулы липидов содержат полярную "голову" и два гидрофобных углеводородных "хвоста". Молекулы ориентированы "хвостами" внутрь мембраны. Такая конструкци оказывается стабильной и энергетически выгодной (по структуре она аналогична пленке мыльного пузыря). Молекулы белков взаимодействуют с такой двухслойной структурой. Белки с гидрофобными поверхностями взаимодействуют с "хвостами" липидов и встраиваются в саму мембрану, а с гидрофильными — соединяются с поверхностью мембраны. По химическому составу мембраны весьма разнообразны. Толщина мембраны около 10 нм. В состав мембраны входит множество механизмов, необходимых для работы клетки. Тут будет рассматриватьcя только те, что имеют отношение к передаче нервных сигналов.
![]() Натриевый насос.
Натриевый насос.
Внутри клетки, в цитоплазме, преобладают ионы K+, снаружи — Na+. Активный мембранный транспорт, натриевый насос, переносит K+ внутрь, а Na+ — наружу. Такой перенос происходит в направлении роста электрохимического потенциала и требует затрат энергии. Для работы натриевого насоса используется энергия гидролиза АТФ (аденозинтрифосфата) — основного энергетического аккумулятора клетки. Механизм переноса использует белокноситель, обозначим его C, образующий комплекс с ионами на одной стороне и отщепляющий эти ионы на противоположной стороне мембраны. Ни носитель, ни комплексы CK, CФNa с ионами Na+, K+: не покидают мембрану. В итоге через мембрану проходят потоки ионов K+ и Na+, направленные: K+ — внутрь клетки, Na+ — наружу.
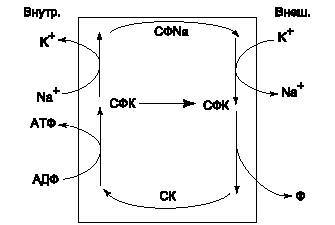 |
| (Рис.5) Термодинамическая схема натриевого насоса. |
В результате работы натриевого насоса концентрация K+ и Na+ становится неравновесной, но электрический потенциал мембраны не меняется, так как количество ионов K+, перенесенных внутрь клетки, совпадает с количеством ионов Na+, перенесенных наружу. Поэтому суммарный перенос заряда равен нулю и электрический потенциал цитоплазмы не меняется.
![]() Калиевые каналы.
Калиевые каналы.
В мембране существуют каналы, пропускающие только ионы K+ в обоих направлениях. Каналы являются управляемыми, и могут открываться или закрываться в зависимости от разности потенциалов между цитоплазмой и внешней средой, а также обладают инерционными свойствами.
В состоянии покоя часть K-каналов открыта. Из-за разницы концентраций K+ и Na+ на разных сторонах мембраны ионы K начинают диффундировать через K-каналы из цитоплазмы наружу. Этот процесс приводит к оттоку положительного заряда из клетки, и цитоплазма заряжается отрицательно. Возникают кулоновские силы, препятствующие дальнейшей диффузии ионов K+. Как только эти процессы уравновешивают друг друга, диффузия ионов K+ через K-каналы прекращается, и потенциал цитоплазмы достигает равновесного состояния -70 мВ.
В состоянии покоя разница потенциалов на мембране определяется формулой Ходжкина-Катца:
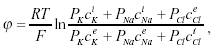 |
Где Pk—проницаемость для
мембраны для ионов K+, Pk—концентрация K+ внутри клетки,
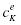 —снаружи,
аналогично для других ионов. Формула выводится, исходя из однородности
поля на мембране, если диффузионные токи IK и INa известны. Значение
потенциала покоя -70..-80 мВ, полученное по этой формуле, согласуется с
экспериментальными значениями.
—снаружи,
аналогично для других ионов. Формула выводится, исходя из однородности
поля на мембране, если диффузионные токи IK и INa известны. Значение
потенциала покоя -70..-80 мВ, полученное по этой формуле, согласуется с
экспериментальными значениями.
Калиевые каналы открываются при изменении потенциала в положительную сторону и остаются открытыми, пока потенциал сохраняет свое значение.
![]() Натриевые каналы.
Натриевые каналы.
Na-каналы аналогичны калиевым, но пропускают только ионы Na+. Отличаются также уровнем потенциала, открывающим канал, и инерционными характеристиками. В состоянии покоя Na-каналы закрыты. Натриевые каналы также открываются при изменении потенциала протоплазмы в положительную сторону. Закрываются сами по себе, через некоторое время после открытия. Закрывшись, находятся в состоянии рефрактерности в течение примерно 1 мс и не могут открыться снова до окончания рефрактерности.
![]() Возникновение нервных импульсов.
Возникновение нервных импульсов.
Механизмы мембраны, ответственные за возникновение нервных импульсов приведены на рис. 6.
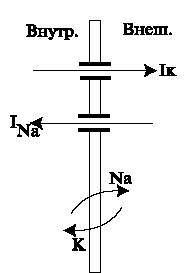 |
| (Рис.6) Механизы мембраны, участвующие в возникновении нервного импульса. |
- Возбуждение импульсов происходит по принципу "все или ничего". Если потенциал цитоплазмы станет положительнее, чем -50 мВ, то произойдет генерация импульса. Изменение потенциала можно проводить микроэлектродом, введенным в цитоплазму и подключенным к источнику напряжения. Деполяризацией мембраны называется изменение ее разности потенциалов от состояния равновесия в положительную сторону, например, от -70 до -50 мВ.
- Если мембрана деполяризуется до уровня -50 мВ, то открываются натриевые каналы, и поток ионов Na+ начинает поступать в клетку. Возникает ток INa, направленный внутрь клетки. Это приводит к дальнейшей деполяризации мембраны. Т.к. Na-каналы управляются разностью потенциалов на мембране, дальнейшая деполяризация открывает все новые Na-каналы, что приводит к росту INa. Положительная обратная связь в данном процессе дает резкий скачок потенциала в сторону деполяризации.
- Натриевые каналы закрываются самопроизвольно, через некоторое время после открытия. Закрывшись, находятся в состоянии рефрактерности около 1 мс и не могут быть открыты вновь, несмотря на деполяризованность мембраны. Ток INa падает до нуля, деполяризаци прекращается. В этот момент потенциал достигает значения +40 мВ.
- Калиевые каналы тоже управляются потенциалом, но более инертны и открываются с задержкой по сравнению с натриевыми. Т.к. потенциал смещен в сторону деполяризации, открытие калиевых каналов приводит в диффузионному калиевому току, направленному из клетки. Ток IK нарастает медленно и направлен противоположно INa. К моменту, когда IK достигает максимума, ток INa уменьшается, и потенциал начинает меняться в противоположную сторону: мембрана реполяризуется.
- За счет тока IK мембрана реполяризуется до исходного потенциала -70 мВ. За счет инерционности K-каналов мембрана гиперполяризуется до -90 мВ. Калиевый ток прекращается.
- По окончании импульса натриевые каналы пребывают в состоянии рефрактерности около 1 мс, когда возникновение нового импульса невозможно. Происходит деполяризация до –70..-80мВ — равновесного мембранного потенциала.
- Под действием изменения потенциала открываются натриевые каналы на соседнем участке, и процесс возбуждения распространяется вдоль волокна.
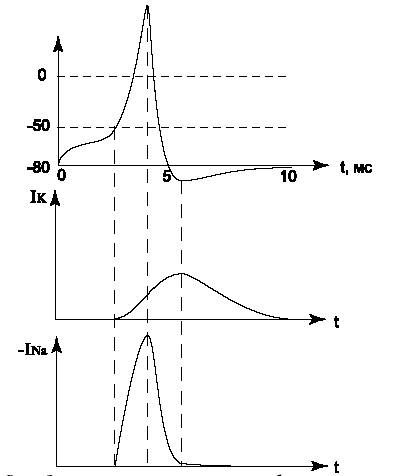 |
| (Рис.7) Зависимость ионных токов и мембранного потенциала от времени. |
В результате прохождения импульса часть ионов калия выходит наружу, а часть ионов натрия входит в клетку, причем в итоге потенциал возвращается к исходному значению –80 мВ. Следовательно, количество ионов Na, вошедших в клетку, в точности равно количеству ионов K, вышедших из клетки. Натриевый насос восстанавливает разницу концентраций очень быстро. Так как емкость мембраны мала (C=1 мкФ/кв.см),то суммарное количество ионов, прошедших через мембрану, невелико. Выход ионов каждого вида при прохождении импульса составляет 12 3,7 10. . моль/см2. Восстановление концентраций требует затрат энергии, которая берется из реакции АТФ -> АДФ.
![]() Сальтаторный механизм распространения
импульса.
Сальтаторный механизм распространения
импульса.
Сальтаторный механизм встречается для миелинизированных волокон. В этом случае участки волокна, покрытые миелином (шванновскими клетками) не проводят ток, зато в неизолированных местах, перехватах Ранвье, плотность ионного тока возрастает в 10 раз по сравнению с волокнами, лишенными миелиновой оболочки.
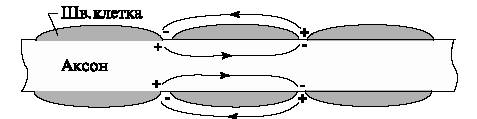 |
| (Рис.8) Сальтаторный механизм распространения. |
Распространение импульса состоит в последовательном возбуждении перехватов Ранвье. Импульс передается "скачками" от одного перехвата к следующему. Так как большая часть волокна изолирована, то суммарный обмен ионами намного меньше, чем для немиелинизированных волокон. Скорость передачи возрастает в 10-50 раз, а энергии расходуется меньше. Дл немиелинизированных волокон кошки скорость передачи — 0,7-2,3 м/с, а дл миелинизированных — 50-100 м/с. Поэтому сальтаторный механизм часто оказывается более эффективным. Такой вид распространения НИ встречаетс только у высших животных.
![]() Синаптическая передача.
Синаптическая передача.
Синапс – соединение для передачи нервного импульса от нейрона к нейрону или от нейрона к мышечному волокну. Синапсы бывают химические и электрические, в центральной нервной системе преобладают химические. В месте контакта мембраны клеток не сливаются, между ними всегда существует небольшой промежуток — синаптическая щель.
Электрический синапс: ширина щели — 2-4 нм (при толщине мембраны 7-10нм). Между контактирующими мембранами образуются две системы каналов: цитоплазма – внешняя среда и цитоплазма – цитоплазма. Первый тип каналов регулирует обмен ионов калия , натрия и хлора. Межклеточные каналы второго типа имеют низкую утечку во внешнюю среду и передают импульсы с использованием того же механизма, что и при передаче импульса по волокну. Задержка передачи для электрических синапсов очень мала. Недостаток электрических синапсов — нерегулируемость: они не реагируют на биологически активные вещества и не меняют свою проводимость. Встречаются у беспозвоночных и низших позвоночных.
В мозгу человека и высших животных преобладают химические синапсы. У одного нейрона бывает от 300 до 20 000 синапсов между аксонами, аксонами – дендритами, дендритами – дендритами, аксонами и мышечными волокнами, аксонами и телом клетки и т.п.
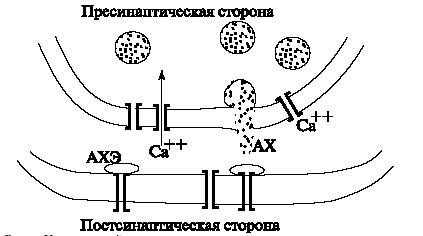 |
| (Рис.9) Химический синапс. |
![]() Порядок синаптической передачи.
Порядок синаптической передачи.
- Импульс подходит к пресинаптической части волокна.
- Открываются кальциевые каналы, ионы Ca++ поступают в пресинаптическую цитоплазму.
- В пресинаптической цитоплазме постоянно находится большое количество синаптических пузырьков — образований, содержащих молекулы вещества — медиатора. Самым распространенным медиатором является ацетилхолин (АХ), кроме него существует около 20 других видов медиаторов. Вследствие появления ионов Ca++ в пресинаптической цитоплазме посредством неизвестного пока механизма пузырьки подходят к мембране и лопаются, сливая медиатор в синаптическую щель. Пузырьки выбрасываются не по одному, а квантами по несколько пузырьков. За один квант в щель попадает 103 – 104 молекул медиатора. За один импульс проходит 100 – 200 квантов медиатора. Даже в отсутствие нервных импульсов каждую секунду выбрасывается несколько квантов медиатора, и синапс поддерживается в состоянии готовности к передаче.
- Попавший в щель медиатор диффундирует на постсинаптическую сторону щели. Этот процесс требует около 0,5мс и вносит существенный вклад в задержку передачи.
- Молекулы медиатора улавливаются рецепторами на постсинаптической стороне:
- Попадание ацетилхолина на рецептор увеличивает проводимость канала в обоих направлениях для ионов Na+ и K+ в равной степени. Это эквивалентно небольшому кратковременному "проколу" мембраны. Ацетилхолин, попавший в рецепторы, гидролизуется ацетилхолинэстеразой (АХЭ), во избежание забивания областей рецепции молекулами АХ. Яд кураре и подобные ему вещества попадают в молекулы АХЭ, предотвращают гидролиз АХ и останавливают работу синапсов.
- Открытие каналов на постсинаптической стороне вызывает поток ионов натрия внутрь, а калия наружу. Возникший ионный ток возбуждает нервный импульс, который распространяется дальше по постсинаптическому волокну.
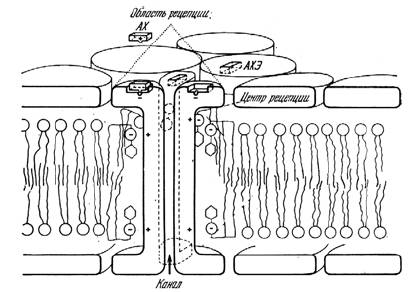 |
| (Рис.10) Химический синапс. |
Существуют медиаторы, которые работают по-другому. Так как контакты бывают дендро-дендритные, дендро-аксональные, то синапсы оказываютс разнообразными по структуре.
Каналы являются специфическими для различных ионов (калия, натрия, хлора). В зависимости от вида ионов, постсинаптическая мембрана или гиперполяризуется при прохождении соответствующих ионов через канал (тормозный синапс), или деполяризуется (возбуждающий синапс).
Синапсы образуются в различных участках клетки. Информационные функции клетки определяются расположением этих участков и их влиянием на мембрану и могут быть очень разными. Например, в зоне аксонного холмика, где обычно генерируется импульс, деполяризация мембраны с наибольшей вероятностью вызовет возникновение нервного импульса.
Отдельный нервный импульс слабо влияет на постсинаптическую клетку и не является самостоятельным носителем информации. Для существенного возбуждения необходимо поступление пачки импульсов. Плотность импульсов в пачке и распределение плотности во времени является носителем информации в нервной системе. Пачка нервных импульсов является аналоговым сигналом, т.к. плотность импульсов в пачке может меняться непрерывно.
Подпороговое возбуждение может распространяться градуально, с затуханием. В коротких дендритах используется этот механизм. Градуальные синаптические потенциалы тоже являются аналоговыми сигналами. Поэтому нервная система человека и животных оказывается аналоговой информационной системой.

![]()
| © Швырев Олег, 2002 | Разработано FiSHES |